
Genome sequencing
We assembled an extensive collection of 49 archaeological grape pips from France (N = 47) and Ibiza (N = 2), all originated from waterlogged environments exhibiting well-preserved morphology and organic material (Fig. 1; Supplementary Data 1). This material spans a broad temporal range from the Bronze Age (2300–2000 BCE, Before Common Era) to the end of the Medieval period (1400–1500 CE) (Fig. 1a). The majority of samples date to the Roman period (50 BCE-500 CE), and provide wide geographic coverage across present-day France (Fig. 1b). Two pips from Ibiza were also included to provide insights on a key trading hub of the ancient Mediterranean world17.
Fig. 1: Archaeological contexts, ancient samples and genomes.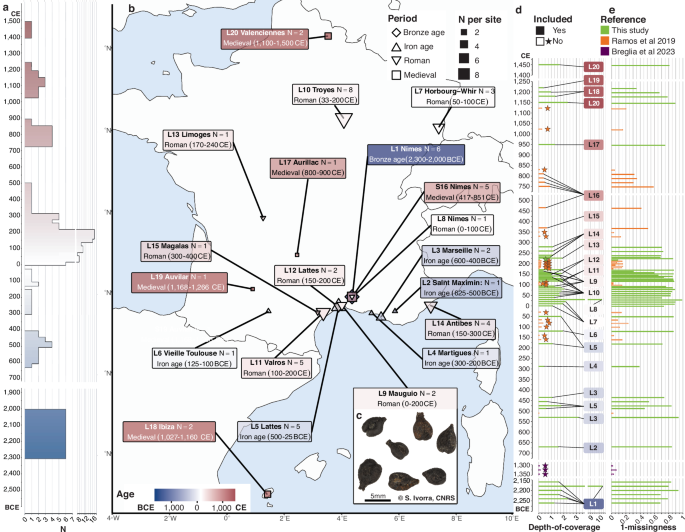
a Temporal range of the 20 archaeological sites investigated. The colour gradient spans the Bronze Age (blue) to the Medieval period (red). b Map of the archaeological sites investigated, with colours reflecting their temporal range. Symbol shapes correspond to the Bronze Age (diamond), the Iron age (triangle), the Roman period (inverted triangle), and the Medieval period (square). Symbol sizes are proportional to the number of ancient grape pips analyzed. An unique label (LX) is assigned to each site and used for referencing throughout the text. c Ancient grape pips from Mauguio (S9, France). d Average depth-of-coverage. Samples sequenced for the first time in this study are coloured in green. One of these newly sequenced samples and most samples from Ramos-Madrigual et al.16 and Breglia et al.19, except six, were excluded due to insufficient coverage (i.e., < 0.1-fold). e Completeness. The values indicate the difference between one and proportion of missing data in the full matrix of 1,045,070 SNPs. All samples considered for downstream analyses show at most 69.64% missingness.
Endogenous DNA content ranged from 3.72 to 66.51% (median = 13.42%), sufficient for cost-effective genome characterization through shotgun sequencing. As expected for ancient DNA, sequencing data showed characteristic patterns of post-mortem DNA damage, including an enrichment of purines at reference positions immediately adjacent to read termini (Supplementary Fig. 1a). Sequencing data also exhibited increasing C > T and G > A mis-incorporation rates toward the end of DNA fragments (Supplementary Fig. 1c; Supplementary Data 3). These signatures are consistent with post-mortem DNA fragmentation (Supplementary Fig. 1d; Supplementary Data 4) primarily driven by depurination, and cytosine deamination reactions concentrated within single stranded overhangs of DNA molecules18. Deamination rates were estimated between 5 and 57 times higher in single-stranded overhangs compared to double-stranded regions (Mean = 17.55; Supplementary Fig. 1e and Supplementary Data 5). Combined, these results support the authenticity of the ancient sequence data generated.
After removing PCR duplicates and low-quality alignments, the average depth-of-coverage across samples was ~1.80-fold (median = 1.60-fold, range = 0.07–10.43-fold, with 39 above 1-fold; Fig. 1d). The newly-generated sequence data were analyzed alongside a comprehensive genome panel of ancient16,19 and modern accessions meeting minimum coverage requirements (Fig. 1e), and capturing global grapevine genetic diversity20,21,22,23,24,25,26,27. Data pseudo-haploidization and conditioning on transition SNPs28 yielded a final data set of 5.79 million SNPs for downstream population genetic analyses, which included 1.05 million with allele frequencies superior to 5%.
The emergence and development of viticulture practices
Principal component analysis (PCA; Fig. 2), phylogenetic relationships and ADMIXTURE29 ancestry profiles (Fig. 3; Supplementary Figs. 2, 3) of modern grapevine accessions reproduced the major genetic clusters previously identified. These included four distinct lineages of Vitis vinifera subsp. sylvestris (hereafter referred to as V. sylvestris): two predominantly found in central (Syl-W1) and western (Syl-W2) Europe, and two others common in the Levant (Syl-E1) and the Caucasus-South Caspian region (Syl-E2) (Fig. 2a). In addition to these wild lineages, six major V. vinifera clusters were also identified, corresponding to western Asian table grapevines (CG1), Caucasus wine grapevines (CG2), muscat grapevines from eastern Asian to western European (CG3), as well as Balkan (CG4), Iberian (CG5) and western European (CG6) wine grapevines (Fig. 2b). ADMIXTURE29 genetic ancestry profiles, together with clustering based on phylogenetic relationships and PCA (Supplementary Figs. 3, 9; Supplementary Data 17, 21), revealed further substructure within the four V. sylvestris lineages and the CG1, CG3, CG4, CG5 and CG6 domesticated clusters, leading to the identification of 20 additional sublineages (Fig. 3a; Supplementary Figs. 4, 7, 10).
Fig. 2: Modern population structure and Principal Component Analysis (PCA).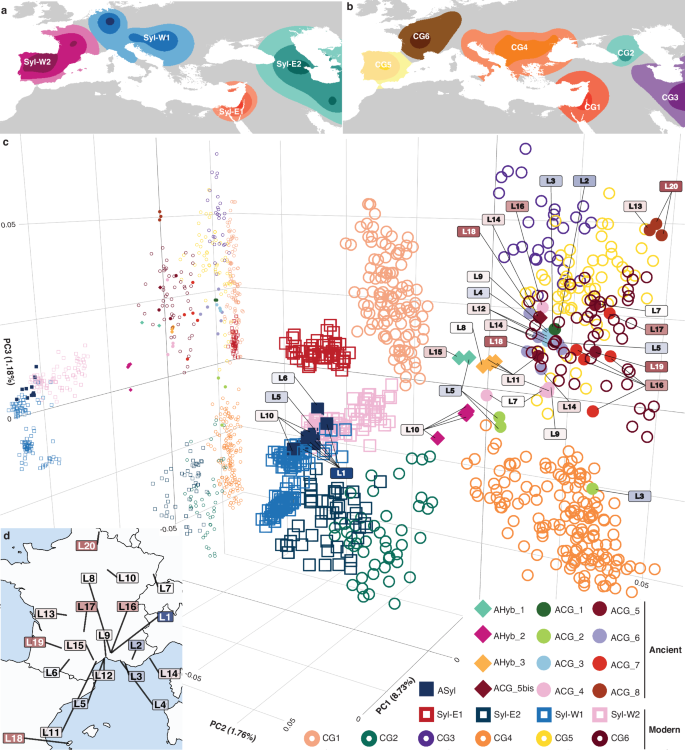
a Map of the main genetic clusters of modern Vitis sylvestris accessions identified in Europe and Southwest Asia. b Same as (a), for modern V. vinifera accessions. The distribution ranges reflect the most frequent accession found in each individual area, as originally described and colour-coded by Dong and colleagues20. For clarity, the geographic extent of these clusters in regions where they are found in lower abundance is not shown. c 3D-PCA projecting ancient samples (n = 54) on the space defined by modern accessions of V. vinifera subspecies (n = 743). The three axes show the first three principal components, with their respective fraction of variance explained between parentheses. 2D-PCA is also shown on the plan formed by PC1 and PC3. Squares, circles and diamonds refer to V. sylvestris, V. vinifera, and their hybrids, respectively. Open and filled symbols indicate modern accessions and ancient samples, respectively. Symbols are coloured according to populations. Simplified labels point to the archaeological sites where samples were excavated, with colours reflecting their age using the same gradient as in Fig. 1a, b. d Summary map of the archaeological sites and their labels.
Fig. 3: ADMIXTURE29 genetic ancestry profiles.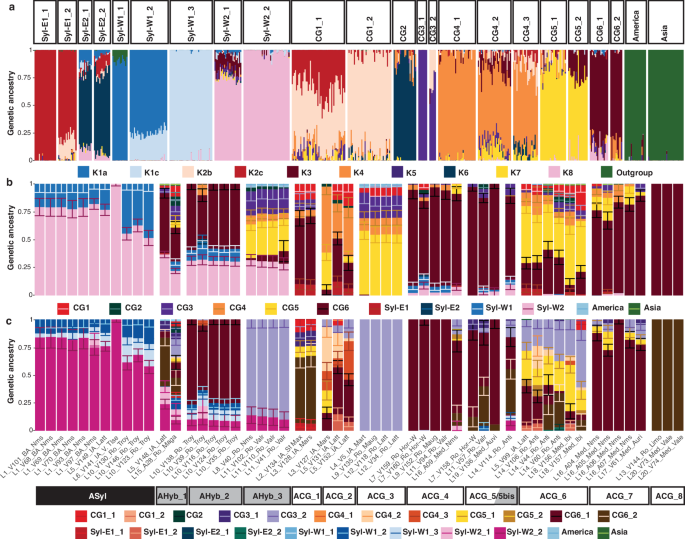
a Unsupervised ADMIXTURE29 of modern accessions (n = 559) considering K = 13 ancestry components. Modern accessions are sorted according to their taxonomic status, with Vitis sylvestris to the left, outgroups to the right, and V. vinifera in the middle. Colours reflect the main genetic ancestry components first characterized by Dong and colleagues20 with minor modifications. K1 ancestry is subdivided into K1a and K1b, which are predominantly found in the Syl-W1_1 and Syl-W1_2 subpopulations, respectively. Similarly, K2 is divided into K2b and K2c, primarily associated with the CG1 and Syl-E1 populations, respectively. Labels shown at the top indicate the subpopulation clustering considered. b Supervised ADMIXTURE29 at the population level of ancient samples with 50 bootstrap pseudo-replicates (n = 54). c Same as (b) at sub-population level. Error bars represent the standard error of the mean (SEM) of ancestry coefficients across individuals, and bars are centred on the mean ancestry coefficient. Error bars are displayed only for ancestries with mean coefficients greater than their SEM. The labels provided within black, grey and white rectangles refer to the various ancient genetic clusters identified among wild, hybrid and domesticated accessions, respectively. The prefix ‘A’ is added to refer to ancient samples and avoid any confusion with those genetic clusters identified from modern accessions. Sample names reflect their original label, followed by their age category (BA = Bronze Age, IA = Iron Age, Ro = Roman, and Med = Medieval), and archaeological site.
PCA projection onto the genetic variation observed in modern accessions revealed that the ancient grape pips spanned a broad range of genetic diversity. While some individuals clustered closely with major wild or domesticated lineages, others occupied intermediate positions, suggesting admixed ancestry (Fig. 2c).This material encompassed the Iron Age (L5, 500–475 BCE) site of Lattes (southern France), and a diversity of Roman sites from both southern (L8 Nîmes, 0–100 CE; L11 Valros, 100–200 CE and L15 Magalas, 300–400 CE) and northern France (L10 Troyes, 33–200 CE).
A total of 11 ancient samples clustered along Principal Component 1 (PC1) with the Syl-W1 and Syl-W2 wild lineages, characteristic of modern western Europe V. sylvestris. This group includes samples from southern France, dating to the Bronze Age (S1 Nîmes, 2300–2000 BCE) and the Iron Age (L5 Lattes, 500–475 BCE; and L6 Vieille-Toulouse, 125–100 BCE), and from Roman northern France (L10 Troyes, 33–85 CE). ADMIXTURE29 analysis revealed mixed ancestry profiles dominated by Syl-W2 (57.3–100.0%), sometimes also incorporating contributions from Syl-W1 (0–42.7%) (Fig. 3c). qpADM30 modelling supported these findings, indicating Syl-W2 as the primary source mixed with Syl-W1 (Syl-W1_2 and Syl-W1_3) in samples from Bronze Age Nîmes (L1), Iron Age Lattes (L5), and Roman Troyes (L10). These results demonstrate that the genetic lineages represented by Syl-W2, today the most prevalent wild grapevine in France, were already established by 2300–2000 BCE. This highlights that western European wild grapes evolved in continuity and the absence of genetic influence from domestic grapevines for over 4000 years of viticulture. The remarkable genetic stability of these wild populations may reflect limited introgression from cultivated varieties, consistent with very low pollen-mediated gene flow from cultivated to wild populations in present-day southern France31. It also aligns with previous genomic work reporting that the vast majority of introgressive events occurred from wild grapevines into cultivated grapes rather than the other way around32. Combined, our findings depict reduced fitness for hybrids in natural forest habitats and/or intentional practices avoiding expanding cultivated forms outside of cultivated environments.
Interestingly, Iron Age sites of southern France also yielded grape pips with genetic compositions characteristic of modern domesticated varieties (Fig. 2c), with ADMIXTURE29 indicating substantial genetic affinities with CG3, CG4, and CG5 in ancient pips from Marseille (V127, 600–400 BCE) and Martigues (V5, 300–200 BCE), as well as mixed profiles dominated by CG6 and eastern ancestries (CG3, Syl-E1 and CG1; Figs. 3b, c). The site of Lattes also yielded grape pips with genetic makeup characteristic of modern domesticated varieties (Fig. 2c), with qpADM30 a modelling (Fig. 4e; Supplementary Fig. 16) and ADMIXTURE29 (Figs. 3b, c) pointing to various combinations of CG3-CG6 genetic affinities (V132, 575-450 BCE; V131, 475–450 BCE; and V99, 125–25 BCE). These findings indicate that Iron Age viticultural practices in southern France involved both the integration of local wild lineages and diverse cultivated varieties. This likely results from multiple intertwined exchanges throughout the Mediterranean, since genetic affinities link to the Levant (CG1-Syl-E1), southwest Asia (CG3), the Balkans (CG4) and Iberia (CG5; Fig. 2b).
Fig. 4: qpADM30 modelling of ancient pips.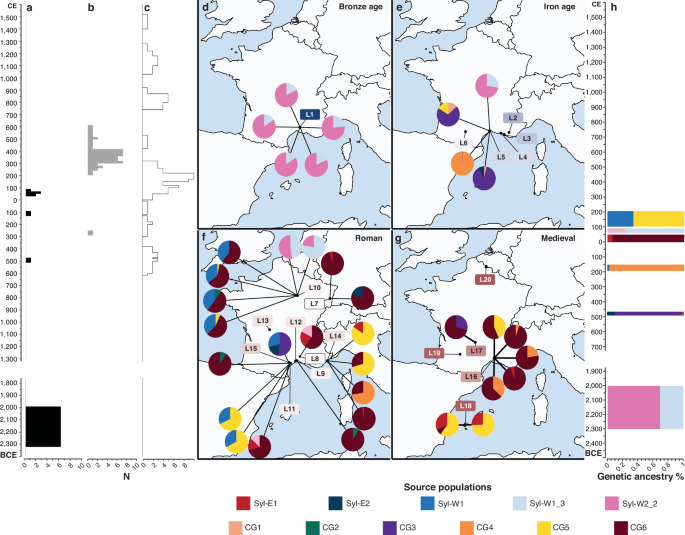
a Temporal distribution of ancient Vitis sylvestris (ASyl). b Same as (a), for ancient hybrids (AHyb). c same as (a), for ancient V. vinifera (ACG). d Genetic makeup of the Bronze Age samples (n = 5). Models were evaluated using a χ²-based goodness-of-fit test as implemented in qpADM30, and only models with associated upper-tailed p ≥ 0.01 are shown. Colours reflect the main source populations identified in models with highest p. e Same as d), for the Iron Age (n = 4). f Same as (d), for the Roman period (n = 19). g Same as (d), for the Medieval period (n = 8). h Earliest temporal occurrence for the diverse genetic sources identified.
Viticultural practices were also conducive of wild admixture since one sample from Lattes (L5: V148, 500–475 BCE) exhibited a mixed genetic makeup including substantial (39.6%) wild ancestry from Syl-W2, with balanced contributions from both Syl-W2-1 and Syl-W2-2 sublineages (Figs. 3b, c). Syl-E1 ancestry, characteristic of modern Levant wild populations, was also found in small proportions in samples from Saint-Maximin (V134, 800–540 BCE), Marseille (V128, 500–400 BCE), and Lattes (V131, 475–450 BCE; V132, 200–150 BCE; and V99, 125–25 BCE) (Figs. 3b, c). This demonstrates that Iron Age viticulture practices of southern France involved admixture both from domesticated grapes and wild grapevines.
The Roman period, which is the most represented in our dataset, reveals a notable shift in the genetic composition of the grape pips investigated. A majority of samples from this period exhibited ancestry profiles dominated by the influence of lineages characteristic of present-day France (CG6) or Spain (CG5) (Fig. 4f; Supplementary Fig. 14c). The CG6 ancestry was found across both southern (Nîmes, 0–100 CE; Mauguio, 0–200 CE; Valros, 100–200 CE; and Antibes, 150–300 CE) and northern regions (Horbour-Whir, 0–50 CE; and Troyes, 33–200 CE). In contrast, CG5 ancestry, while present at Troyes (100–200 CE), appeared more limited in the north, accounting for only 2.4–5.9% of the genetic makeup in two samples (V35 and V124, 100–200 CE), according to qpADM30 modelling. By comparison, CG5 contributed at least 67.7% genetic ancestry in samples from the south (Valros, V102-V110, 100–200 CE; and Antibes, V106-V44, 150–300 CE) (Fig. 4f). This pattern, although enhanced, was confirmed by ADMIXTURE29, which showed relatively widespread CG6 ancestry, and CG5 ancestry confined to southern France (Fig. 3b; Supplementary Figs. 14, 16). CG1 ancestry, which is associated with cultivars from the Levant (Fig. 2b), was also present both in the south (Mauguio, V135, 0–200 CE; Lattes, V118–V34, 150–200 CE; Antibes, V106-V60, 150–300 CE; and Magalas, A28_R, 300–400 CE), and in the north (Horbourg-Whir, V160, 0–50 CE) (Supplementary Fig. 14c).
The Roman period also saw the continuation of viticultural practices involving the mixing of local wild grapevines with domesticated varieties, a trend already observed during the Iron Age. Substantial Syl-W1 ancestry (≥32.2%) was detected in samples from both northern and southern sites. The Syl-W2 lineage also contributed significantly in the south, with 12.6-16.9% ancestry according to qpADM30 (Fig. 4f). ADMIXTURE29 results indicate that Sylv-W2 ancestry was also present in northern regions, particularly in Troyes, where more than half of the detected wild introgressions could be attributed to Syl-W2 (Supplementary Figs. 14c, 15c).
Furthermore, Syl-E1 ancestry, typical of wild grapevines from the Levant, was identified in one sample from Horbourg-Whir (0–50 CE), as well as in southern samples from Nîmes (0–100 CE), Valros (100–200 CE) and Antibes (150–300 CE) (Fig. 4f, Supplementary Figs. 14c, 15c). These findings suggest the influence of non-local wild grapevines into cultivation. Similarly, Syl-E2 ancestry, characteristic of present-day wild grapevines from the Caucasus (Fig. 2a), was detected in samples from Horbourg-Whir (0–50 CE), Magalas (300–400 CE) (Fig. 4f; Supplementary Fig. 14c), Nîmes (0–100 CE),Valros (100–200 CE) and Antibes (150–300 CE) (Supplementary Fig. 14c). CG2 ancestry–a lineage most commonly found today in cultivars from the Caucasus (Fig. 2b)–was also detected in two samples from southern France (Mauguio, 0–200 CE; Valros, 100–200 CE) and in one sample from Troyes (33–65 CE), according to qpADM30 (Fig. 4f). Combined, these findings point to continuing long-distance contacts during the Roman period, with seeds or cuttings likely transported, either directly from the Levant and the Caucasus, or indirectly through Greece or Italy, into France. These introductions appeared to have been successfully integrated into local viticultural systems through sexual reproduction, contributing to the genetic diversity of cultivated grapevines in Roman Gaul.
Importantly, the genetic composition of Medieval grapes closely resembled that observed during the Roman period (Fig. 4g). The two major domesticated lineages found in present-day France and Spain (CG6 and CG5) dominated the ancestry of all individuals that could be successfully modelled using qpADM30. ADMIXTURE29 also revealed the widespread presence of CG6 ancestry in France and Ibiza, although CG5 ancestry predominated in Ibiza and southern France, aligning with alternative significant qpADM30 models describing pips from Nîmes (A09, 417–515 CE; A04 and A07, 731–851 CE) as CG6 and CG5 mixtures (Supplementary Fig. 16). Although qpADM30 modelling failed for all individuals from Valenciennes in northern France, ADMIXTURE analyses29 revealed that two individuals (V73, 1100–1200 CE; and V74, 1400–1500 CE) carried ancestry mainly derived from the CG6 lineage, specifically the CG6_2 sublineage (Fig. 3b, c). At the same time, qpADM30 models show substantial influences from non-local lineages, including wild ancestries such as Syl-E1 (5.7% in one sample from Nîmes, A07_M, 731–851 CE; and 25.8–32.3% in two samples from Ibiza, V156 and V157, 1027–1160 CE), and domesticated components such as CG3 (28.9% in one sample from Aurillac, V61, 800–900 CE), which is characteristic of the Levant and Central Asia (Fig. 4g). Similar patterns were observed in ADMIXTURE analyses29, with CG3 ancestry detected not only in Medieval Aurillac, but also in Medieval Ibiza and Nîmes. These findings suggest that oriental influences persisted into the Medieval period, with continued integration of non-local genetic material into local grapevine populations.
Finally, samples identified as carrying pure wild ancestries displayed a distinct temporal distribution compared to those with purely domesticated or hybrid ancestries (Fig. 4a–c; two-sided Mann–Whitney U test p < 1.19e–3). Samples with exclusively wild genetic profiles were indeed present from the Bronze Age (2300–2000 BCE) to the Roman period (60–85 CE). In contrast, hybrid samples showing mixtures of wild and domesticated ancestries were first detected in the mid-Iron Age (500–475 BCE), and persisted into the Roman period (300–400 CE). Samples with genetic profiles consisting of ancestries representative of modern domesticated lineages appeared from the Iron Age (625–500 BCE) to the late Medieval period (1400–1500 CE). This temporal pattern supports the emergence of viticulture in the region between 2000 BCE and 500 BCE, consistent with archaeological evidence pointing to the introduction of grapevine cultivation in southern France during the early 6th century BCE4,5,11. Over time, agricultural practices increasingly favoured the propagation of grapevines with genetic composition strictly combining ancestries found in modern cultivars at the expense of wild grapevines.
Vegetative propagation
Grapes can be propagated either sexually, through seeds, or vegetatively, through cuttings that produce genetically identical clones. To investigate the emergence and extent of vegetative propagation, we searched for genetically identical or closely related samples in our data set using READv233, a methodology originally developed to assess relatedness in ancient human genomes33. We first calibrated the approach through simulations-based analyses using modern grapevine accessions with known pedigrees, establishing normalization thresholds for pairwise genetic distances that minimize false assignment rates (see “Methods”). The calibrated thresholds produced a very low false positive rate, with only 0.4% of unrelated individuals misclassified as related up to the second degree (Supplementary Fig. 17a). They also reliably identified identical domesticated clones and identical wild clones in respectively 99.3% and 94.60% of cases (Supplementary Figs. 17e, 18d). No unrelated domesticated individuals, nor closely related individuals, were identified as identical, confirming the robustness of our approach to identify grapevines that are maintained through vegetative propagation (Supplementary Figs. 17a–d). The method also showed good sensitivity for close kin, correctly identifying 86.9% of first-degree relatives as related up to the third degree (Supplementary Fig. 17d). In contrast, accuracy decreased for identifying second- and third-degree relatives, with 69.8% and 93.0% of such pairs incorrectly classified as unrelated, respectively (Supplementary Figs. 17b, c). To maintain high confidence in our inferences, we therefore restricted our analyses to clone detection and first- or second-degree relatives only, excluding more distant assignments. Due to the overlap in misclassification rates between first- and second-degree relatives (48.8% and 1.7%, respectively), we also grouped these two categories into a single class of closely related individuals.
We first screened for pairs of clones among the ancient grape pips. Strikingly, a substantial fraction of the wild Bronze Age grape pips from Nîmes were genetically identical (Fig. 5a). This finding is surprising, as vegetative propagation is limited in wild grapevines. Given that these pips were recovered in close spatial proximity, in a palaeochannel where seeds may have been deposited naturally through water or gravity, we interpret them as likely deriving from a single grapevine.
Fig. 5: f3-outgroup statistics63 and genetic relatedness between pairs of ancient samples.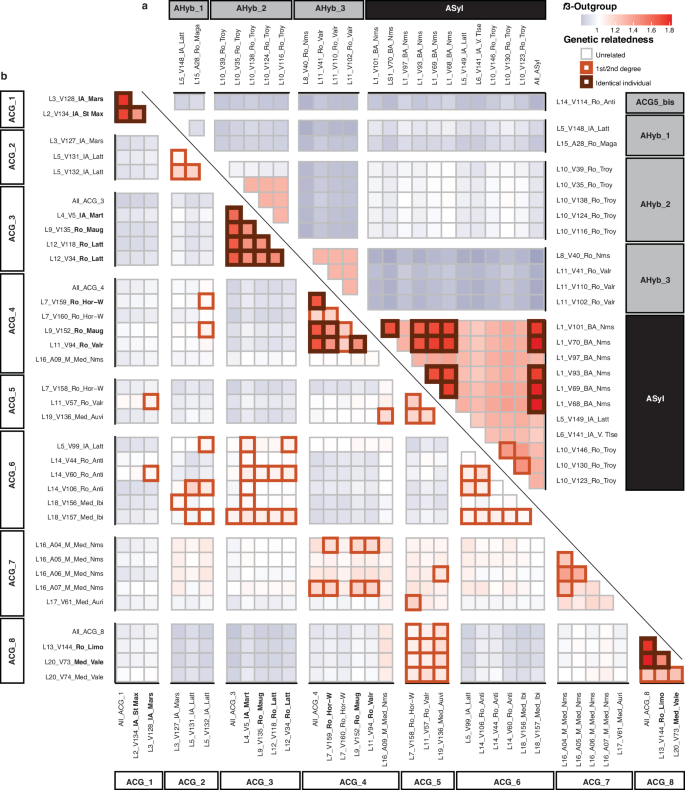
a Ancient Vitis sylvestris and/or ancient hybrids. Samples are grouped according to their genetic clusters. f3-ougroup statistics, using non-V. vinifera species as outgroups, measure the extent of shared genetic drift between pairs of samples. The colour gradient reflects lower (blue) to higher (red) shared genetic affinities. Squares of wider stroke indicate closer genetic relatedness up to the second degree. b Same as (a), for ancient V. vinifera. The labels provided within black, grey and white rectangles refer to the various genetic clusters identified among wild, hybrid and domesticated accessions, respectively. The prefix ‘A’ is added to refer to ancient samples and avoid any confusion with those genetic clusters identified from modern accessions. Groups identified with a “hyb_” prefix correspond to the data merger from several samples identified as genetically identical.
More remarkably, we identified genetically identical clones across distinct archaeological sites dating to roughly similar time periods, suggesting the use of vegetative propagation and the intentional movement of cuttings or, less likely, pips–possibly through commercial exchanges, such as the trade of fruits or preparations in amphorae (Fig. 5b). For instance, clonal samples were found in Iron Age contexts at Saint-Maximin (625-500 BCE) and Marseille (V128, 500–400 BCE), two sites located in southern France and separated by ~40 km. In the 6th c. BCE, Saint Maximin presented all the characteristics of local indigenous settlements found in the region. It especially stood out from those sites found on the outskirts of Marseille, a large neighbouring city with intense seaborne connection with the Greek world. The genetic proximity documented may therefore represent the earliest direct evidence of vegetative material and viticultural knowledge exchanged between coastal trade hubs and rural inland sites. Another case, from the Roman period, involves a pip from Horbourg-Whir (V159, 0–50 CE) in northeastern France, which was genetically identical to two others pips from the South of France (Mauguio, V152, 0–200 CE; and Valros, V94, 100–200 CE), located 560 and 610 km away, respectively. Evidence of long-term vegetative reproduction was also detected across both time and space. The identification of identical clones in two sites separated by a time gap necessarily implies the practice of vegetative propagation, as maintaining self-pollination in hermaphrodite cultivated plants through centuries appears especially difficult. In southern France, identical samples were recovered from Iron Age Martigues (V5, 300–200 BCE) and Roman-period Mauguio (V135, 0–200 CE) and Lattes (V118–V34, 150–200 CE). In central and northern France, clonal continuity was observed over a thousand years between Roman-era Limoges (V144, 170–240 CE) and Medieval Valenciennes (V73, 1100–1200 CE), two sites located approximately 530 kilometres apart. Combined, these findings indicate that vegetative reproduction was a widespread and consistent feature of grapevine cultivation across the entire region investigated from at least the Iron Age (625–500 BCE), with some grapevine clones persisting over one millennium. This enduring clonal continuity echoes historical and archaeological evidence showing the prevalence of layering techniques for propagating grapevines in southern Gaul and the wider Roman world34,35.
Wild grape pips exhibiting first/second degree genetic relatedness were only detected within the same archaeological sites (Fig. 6a). In contrast, such close genetic relationships were identified across both space and time among grape pips showing a domesticated genetic makeup (Fig. 6b). In the southern regions, these relationships occurred over short distances ( < 100 km) as well as long distances ( > 700 km), extending beyond present-day France–for example, between the French Mediterranean coast and Ibiza (Spain). Notably, close genetic relationships also reached further north, connecting Roman Horbourg-Whir to Medieval Valenciennes. The chronological context of each site provided a unique opportunity to trace the cultivation history of individual grapevines across the country. For instance, sample V5 from Iron Age Martigues (300–200 BCE) showed close genetic ties to three samples from the Roman site of Antibes (V60, V106, and V156, 150–300 CE), and two samples from Medieval Ibiza (V156 and V157, 1027–1160 CE). Given that such genetic proximity indicates first- or second-degree relatedness, this pattern suggests that these grapevines underwent no more than four cycles of sexual reproduction over the course of more than a millennium. This aligns with the picture emerging from genetically identical clones as well as archaeological and historical evidence for organized vineyard management, and the wide circulation of cultivars across the western Mediterranean at the time12,13. Combined, these findings support that vegetative propagation was a common viticulture practice during Roman and Medieval periods, highlighting the role of extensive exchange networks in disseminating grapevine cuttings through space and time.
Fig. 6: Genetic relatedness.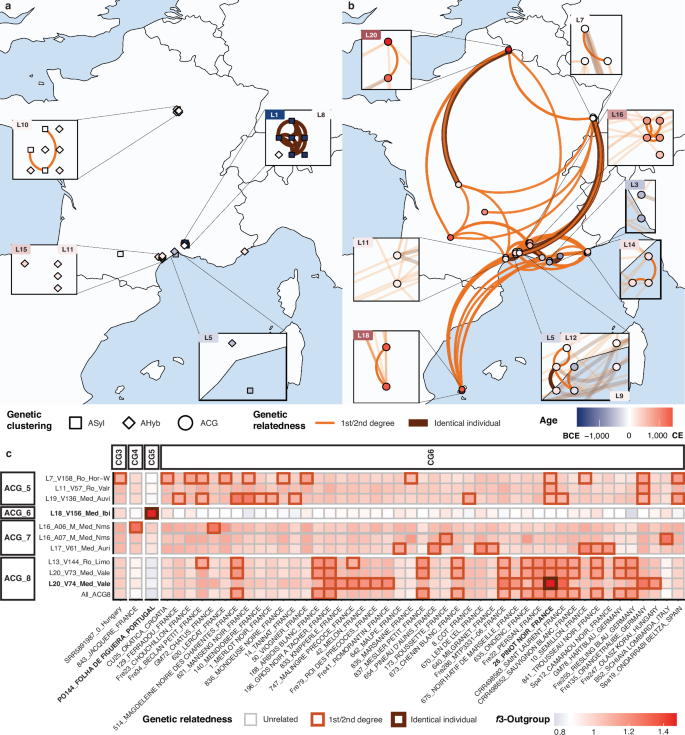
a Amongst ancient Vitis sylvestris and hybrid samples. Contemporary archaeological sites are zoomed in, whether or not they comprised samples genetically related up to the 2nd degree. Transparent lines within the zoomed-in areas indicate the genetic relatedness between the samples from the highlighted sites and those from other locations. b Same as (a), for ancient V. vinifera samples. The colour gradient spans the Bronze Age (blue) to the Medieval period (red), following Fig. 1a, b, and site labels follow Fig. 2d. c Between pairs of ancient samples (vertical axis), and modern accessions (horizontal axis). Samples are grouped according to their genetic clusters. f3-outgroup statistics, using non-V. vinifera taxa as outgroups, measure the extent of shared genetic drift between pairs of samples. The colour gradient reflects lower (blue) to higher (red) shared genetic affinities. The figure only shows those pairs showing at least one genetic relatedness up to the 2nd degree. Lines with a wider stroke indicate closer genetic relatedness.
Given the long-term maintenance of certain cultivars through vegetative propagation, we next examined our dataset for instances of clonal matches or close genetic relatives between ancient samples and modern accessions (Fig. 6c). Strikingly, two Medieval samples were found to be genetically identical to modern cultivars. Sample V156 from Medieval Ibiza (1027–1160 CE) matched with the ‘Folha de Figueira’ cultivar, a white grape variety still grown in Portugal today. Sample V74 from Medieval Valenciennes (1400–1500 CE) was also identical to the renowned red grape variety ‘Pinot Noir’–one of the oldest known cultivated grapevines, believed to have originated in Burgundy, northern France. Our results indicate that ‘Pinot Noir’, a grape variety critical for the global wine industry at the origin of some of the world’s most prestigious wines, was genetically already fully established no later than the end of the 15th century CE. This variety also had a massive influence on modern viticulture, with many cultivars representing first- or second-degree relatives (Fig. 6c).
Dining and Cooking