

Data presented in Table 2 indicates a significant (p < 0.05) increase in ‘Picual’ shoot length rate, the number of new shoots per shoot, and the length of new shoots in response to NPK nano-fertilizers (NPKNF). The application of NPK at 4000 mg L−1 resulted in the highest values in shoot length increase rate (36.31%), number of new shoots per shoot (5.09), and new shoot length (6.04 cm). In contrast, the control treatment exhibited the lowest values for these parameters, with a shoot length increase rate of 24.85%, number of new shoots per shoot of 1.03, and a new shoot length of 3.04 cm. Additionally, all NPKNF treatments significantly (p < 0.05) increased leaf fresh and dry weights compared to the control. The 3000 and 4000 mg L−1 NPKNF treatments yielded the highest fresh weight (0.113 and 0.123 g, respectively) and dry weight (0.067 and 0.070 g, respectively), whereas the control treatment showed the lowest significant (p < 0.05) values for both fresh (0.087 g) and dry weight (0.047 g)., the leaf area was also positively influenced by the NPKNF treatments. The largest leaf area (3.88 cm2) was observed with the 4000 mg L−1 NPKNF treatment, while the control exhibited the smallest leaf area (3.23 cm2).
The observed increment in olive vegetative growth aligns with previous findings on date palm34,35, olive seedlings49,50,67, and Valencia orange33, which also reported significant improvements in vegetative growth following the application of NPKNF. The notable increase in growth characteristics following the application of NPK nano-fertilizers can be attributed to their unique properties, which facilitate deeper penetration into leaf tissues and promote efficient nutrient transport to the plant’s growth centers68,69,70. As a result, nutrient uptake is improved, leading to enhanced plant growth and development.
Table 2 Effect of spraying NPK nano-fertilizers on the vegetative growth characteristics of ‘Picual’ olive trees during the “on-year” season.
The role of NPK nutrients in promoting vegetative growth is well documented. Nitrogen (N) enhances photosynthesis, cell division, and elongation while also increasing meristematic activity by contributing to the synthesis of DNA, RNA, chlorophyll, amino acids, plant hormones (IAA), enzymes, and vitamins8,29,69,71. In addition, phosphorus (P) plays a crucial role in phospholipid synthesis, nucleic acids, and energy compounds (ATP), which enhance photosynthesis and respiration, ultimately leading to greater vegetative growth29,68,69,71,72. Furthermore, potassium (K) contributes significantly by regulating osmotic pressure, activating enzymes related to photosynthesis, and enhancing nutrient uptake, all of which improve plant vigor29,68,69,71. These combined effects of N, P, and K ensure an optimal balance of nutrients, leading to increased shoot length, number of new shoots, and enhanced leaf development.
The fresh and dry weight results of leaf agree with Hagagg et al.67who reported a significant impact of NPKNF on olive seedlings. The highest dry matter accumulation in treated olive leaves is associated with increased photosynthetic pigments, photosynthesis rates, and carbohydrate accumulation, which contribute to overall dry matter content73. This increased dry matter content enhances the plant’s ability to store and utilize essential resources, supporting overall productivity. Similarly, the increase in leaf area due to NPKNF application is consistent with the findings of Roshdy and Refaai34 on date palm, Mhawesh and Mohsen50 on olive seedlings, and El-Shereif et al.33 on Valencia orange. These studies demonstrated that NPKNF significantly enhances leaf expansion, thereby improving photosynthesis efficiency, which further contributes to increased vegetative growth. Table 3 presents data indicating that NPKNF treatments significantly (p < 0.05) enhanced leaf nitrogen, phosphorus, and potassium concentrations compared to the control. Among the tested treatments, the application of NPKNF at a concentration of 4000 mg L−1 resulted in the highest recorded levels of leaf nitrogen (2.85%), phosphorus (0.44%), and potassium (1.44%).
In contrast, the control exhibited the lowest concentrations, with values of 2.28% for nitrogen, 0.016% for phosphorus, and 0.80% for potassium. Similarly, the data in Table 3 demonstrate that NPKNF treatments had a favourable impact on the nutrient composition of the fruit flesh. The highest nitrogen (0.84%), phosphorus (0.36%), and potassium (2.93%) concentrations in the flesh were observed in response to NPKNF application at 4000 mg L−1. Conversely, the control treatment resulted in the lowest recorded values, with nitrogen, phosphorus, and potassium concentrations of 0.73%, 0.24%, and 2.29%, respectively. Such improvements in leaf nutrient composition following NPKNF application have been observed in various horticultural species, including date palm (Phoenix dactylifera)34,35, mango (Mangifera indica)37,38,39, and orange (Citrus sinensis)33. The observed increase in leaf mineral content may be attributed to the role of nanoelements in enhancing vegetative growth and metabolic activity, thereby promoting the accumulation of essential chemical constituents in leaf tissues69,74.
Table 3 Effect of spraying NPK nano-fertilizers on leaf and fruit N, P, and K content of ‘Picual’ olive trees during the “on-year” season.
The data presented in Table 4 demonstrate that fruit weight increased significantly (p < 0.05) in response to the foliar applications of NPKNF compared to the control. The highest fruit weight was recorded in trees treated with NPKNF at 4000 mg L−1 (6.94 g), followed by those treated with NPKNF at 3000 mg L−1 (6.47 g). The lowest fruit weight was observed in the control treatment (5.34 g). Additionally, all NPKNF treatments had a statistically significant impact on fruit volume. Trees sprayed with NPKNF at 4000 mg L−1 exhibited the highest fruit volume (6.09 cm2), followed by those treated with NPKNF at 3000 mg L−1 (5.40 cm2), whereas the control treatment resulted in the lowest recorded fruit volume (4.12 cm2). In contrast to fruit weight and volume, flesh thickness did not exhibit significant (p < 0.05) differences among treatments, as indicated by the data in Table 4. However, flesh weight was notably influenced by NPKNF application, with 4000 mg L−1 treatment yielding the highest recorded value (6.11 g), while the control treatment resulted in the lowest flesh weight (4.52 g). Regarding seed weight, no significant (p < 0.05) differences were observed among the treatments. Nevertheless, the pulp-to-seed ratio was significantly (p < 0.05) affected by the applied treatments, with the highest ratio recorded in the 4000 mg L−1 NPKNF treatment (7.56), whereas the control treatment exhibited the lowest ratio (5.52). The findings of the present study regarding fruit weight align with those of El-Shereif et al.33, who observed that NPKNF application enhanced fruit weight in Valencia orange.
Table 4 Effect of spraying NPK nano-fertilizers on physical fruit characteristics of ‘Picual’ olive trees during the “on-year” season.
Similarly, Gad et al.75 demonstrated that foliar spraying with nano-potassium silicate significantly increased fruit weight in the ‘Ewais’ mango cultivar compared to the untreated control. Roshdy and Refaai34 also reported that NPKNF application enhanced the average fruit weight of date palm. The significant increase in fruit volume observed in our study aligns with the findings of Morales-Sillero et al.76, who reported that NPK fertilizer application improved fruit volume in mature ‘Manzanillo’ olive cultivar. Inglese et al.77 noted that nitrogen and potassium fertilization during the pit-hardening stage increased fruit volume. Our results on flesh weight corroborate those of Abbasi et al.78, who found that combined foliar application of micro- and macronutrients significantly improved olive flesh weight.
Furthermore, the present findings regarding the pulp-to-seed ratio are consistent with Roshdy and Refaai34, who reported that NPKNF significantly increased the pulp percentage in date palm. Ben Mimoun et al.79 also demonstrated that potassium fertilization improved yield and fruit quality, including fruit weight and the flesh (pulp)-to-seed ratio. Several other studies have similarly reported that NPK fertilization enhances both fruit weight and the pulp-to-seed ratio76,77,78,80. The use of nano-based NPK fertilizers has been shown to significantly improve fruit tree productivity75,81. This improvement in fruit characteristics may be attributed to the ability of nano-fertilizers to boost photosynthetic efficiency, thereby increasing photoassimilate production and their subsequent translocation and accumulation in fruits82,83. Additionally, elevated nutrient concentrations in leaves may substantially influence fruit growth and overall yield75,84.
The data in Table 5 indicate that the applied treatments had no statistically significant effect on fruit length, diameter, or shape index. Similarly, yield measurements showed no significant differences between NPKNF-treated trees and the control.
Table 5 Effect of spraying NPK nano-fertilizers on physical fruit characteristics and yield of ‘Picual’ olive trees during the “on-year” season.
The maturity index of ‘Picual’ olive fruits was determined based on skin and flesh colour, following the methodology described in the materials and methods section, using 100 fruits per treatment. Supplementary Figure S1 indicates that fertilization with the NPKNF delayed fruit ripening. These findings are consistent with those reported by Rodrigues et al.85. However, they contrast with the results of Rosati et al.86who concluded that fertilization did not affect fruit maturation.
The moisture and oil content (%) of ‘Picual’ olive fruits were influenced by NPKNF fertilization (Fig. 2). The results indicated that increasing the concentration of the NPKNF led to a corresponding increase in both oil and moisture content compared to unfertilized fruits (control). The oil content (on a dry weight basis) increased from 30.8% in unfertilized olive fruits to 38.9% in fruits treated with the NPKNF at a concentration of 4000 mg L−1.
These findings align with those of Rosati et al.86, who reported that foliar fertilization with potassium and nitrogen enhances oil and moisture content in olive fruits. Fertilization typically increases both the pulp-to-stone ratio and oil content. Since the pulp contains significantly more oil than the stone, an increase in the pulp-to-stone ratio leads to higher overall oil content without altering the oil concentration within the pulp itself. Consequently, the commonly reported increase in oil content associated with fertilization can be attributed to the enhanced pulp-to-stone ratio rather than a direct increase in the oil concentration of the pulp86.
Fig. 2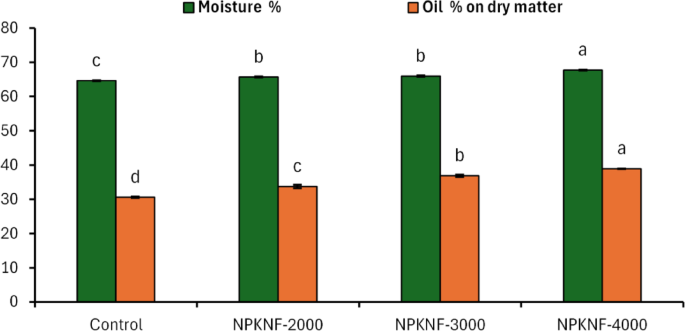
Effect of spraying NPK nano-fertilizers on moisture and oil % of ‘Picual’ olive trees during the “on-year” season. Identical letters within the same trait indicate the absence of statistically significant differences between treatments at the 5% significance level. The error bar represents the standard error (± SE).
Olive oil classification (extra virgin, virgin, ordinary, and lampante) is determined through chemical analysis of free fatty acids (FFA), peroxide value (PV), ultraviolet absorption coefficients (K222 and K270), and ΔK, along with sensory evaluation58. As shown in Table 6, the FFA values ranged from 0.08 to 0.11% as oleic acid, while PV varied between 5.03 and 5.85 meq O₂/kg oil. The ultraviolet absorption values at K222 and K270 ranged from 0.913 to 1.049 and from 0.055 to 0.064, respectively. Importantly, all oil samples, including those from both treated and untreated fruits, exhibited FFA, PV and UV absorbance (at 232 and 270 nm) and ∆K values that were below the maximum limits established by the International Olive Council (IOC) for extra virgin olive oil58. These results align with the findings of Zipori et al.83, Busso et al.87, Tekaya et al.88, and, who reported that while proper fertilization improves tree physiology, photosynthesis, and productivity, its influence on oil quality parameters appears limited. The saponification values, which reflect average fatty acid chain length, were determined to be 193.27 mg KOH/g oil for the untreated control, and 193.22, 193.26, and 193.33 mg KOH/g oil for oils from fruits treated with NPKNF at concentrations of 2000, 3000, and 4000 mg L−1, respectively, all of which comply with both Codex89 and Egyptian90 standards. Furthermore, the phenolic content analysis revealed that foliar fertilization resulted in reduced total polyphenol content (expressed as caffeic acid) compared to the control (350.51 mg/kg), with values decreasing to 298.59, 289.28, and 277.11 mg/kg for oils from fruits treated with 2000, 3000, and 4000 mg L−1 NPKNF, respectively. Since phenolic compounds significantly contribute to olive oil’s nutritional value, sensory characteristics, and oxidative stability91,92,93,94,95, this observed reduction in polyphenol content aligns with findings from previous studies94,96.
The observed reduction in phenolic content following fertilization may result from altered activity of key enzymes including phenylalanine ammonia-lyase (PAL), polyphenol oxidase (PPO), and peroxidase (POD), which regulate phenolic compound biosynthesis. PAL plays a crucial role in this process by catalysing the conversion of phenylalanine to cinnamic acid, thereby directing carbon flow from primary to secondary metabolism through the phenylpropanoid pathway. This enzymatic reaction also serves an important function in nitrogen metabolism by liberating nitrogen from phenylalanine during its transformation to cinnamic acid, facilitating nitrogen redistribution particularly under nitrogen-limited conditions. Following nitrogen fertilization, competition may arise between protein synthesis and phenolic compound biosynthesis for their common precursor, phenylalanine. In such cases, phenylalanine is likely preferentially allocated to protein synthesis rather than being diverted into the phenylpropanoid pathway via PAL activity, consequently restricting phenylpropanoid biosynthesis. This metabolic competition provides a plausible explanation for the decreased phenolic content observed with nitrogen fertilization95,96. Supporting these biochemical observations, antioxidant activity assessments using the 2,2-diphenyl-1-picrylhydrazyl (DPPH) free radical assay revealed that phenolic extracts from oil obtained from unfertilized fruits exhibited superior antioxidant capacity compared to those derived from NPKNF-treated olives, as detailed in Table 6. The observed antioxidant activity demonstrates a positive correlation with total phenolic content91,92consistent with established biochemical relationships. Unsaponifiable matter (UNSAP), comprising hydrocarbons, long-chain alcohols, fat-soluble vitamins, and phytosterols, represents the oil fraction resistant to alkaline saponification but soluble in nonpolar solvents. This component serves as a critical marker for olive oil authenticity verification58. All analysed oil samples showed UNSAP percentages below the International Olive Council’s 1.5% threshold58, with values progressively increasing alongside NPKNF concentration and peaking in the 4000 mg L−1 treatment group. Our findings align with those of Rosati et al.86, who demonstrated that foliar nitrogen fertilization enhances the nutritional and health value of olive oil by increasing the levels of bioactive compounds, such as phytosterols. However, contrasting findings by Jordão et al.97 showed no significant impact of combined nitrogen, phosphorus, and calcium foliar applications on phytosterol content.
Table 6 Quality characteristics of ‘picual’ Olive oil in response to foliar fertilization with a nano mixture of NPK during the “on-year” season.
Values are presented as mean ± standard error (SE). Within each row, values followed by a common lowercase letter are not significantly different (Duncan’s Multiple Range Test, p < 0.05).
Pigment composition substantially affects olive oil quality, influencing both visual characteristics (a primary consumer evaluation criterion) and oxidative stability. Chlorophylls and carotenoids, the primary pigments, measured 1.80 and 1.53 mg/kg, respectively, in control samples, increasing dose-dependently with NPKNF treatment to maximum concentrations of 2.39 and 1.89 mg/kg at 4000 mg L−1.
Chromatographic analysis (Table 7) revealed a characteristic olive oil fatty acid profile spanning C14 to C22 chains, with oleic acid (C18:1) as the predominant component. The chromatographic analysis revealed significant treatment effects on fatty acid composition. Oleic acid (C18:1) content showed a concentration-dependent increase with NPKNF application, rising from 63.50% in the control to 63.83%, 63.89%, and 64.06% at 2000, 3000, and 4000 mg L−1, respectively.
Table 7 Relative percentage of fatty acids of ‘Picual’ olive oil in response to foliar fertilization with a nano mixture of NPK during the “on-year” season.
Conversely, the linolenic acid (C18:3) content decreased progressively with fertilization intensity, measuring 1.00%, 0.93%, and 0.90% in treated samples (2000, 3000, and 4000 mg L−1, respectively) compared to 1.07% in the control. Notably, the control value exceeded the International Olive Council’s maximum limit of 1.0% for linolenic acid58. The NPKNF treatment elevated the total monounsaturated fatty acid (MUFA) content, primarily by increasing oleic acid (C18:1) concentration. Conversely, it reduced the total polyunsaturated fatty acid (PUFA) content, specifically diminishing linoleic (C18:2) and linolenic (C18:3) acids. This shift resulted in a higher MUFA/PUFA ratio than in the control. This rebalancing towards a more monounsaturated profile is known to confer greater resistance to oxidative degradation and rancidity. Calculated iodine values, reflecting the oil’s degree of unsaturation, ranged from 80.28 to 81.04 g iodine/100 g oil across all samples. These variations in iodine value directly correlate with the observed changes in unsaturated fatty acids, particularly oleic (C18:1) and linoleic (C18:2) acids, demonstrating the influence of foliar fertilization on the oil’s fatty acid profile. The sensory profile of olive oil comprises both positive and negative attributes as defined by IOC58. Positive characteristics include fruitiness, bitterness, and pungency, while negative attributes or defects encompass musty, fusty, winey, rancid, metallic, and muddy sediment notes. Phenolic compounds significantly influence this sensory profile, particularly by enhancing bitterness and pungency91,92,93. As presented in Fig. 3, fertilization treatment did not significantly affect fruitiness intensity. However, the unfertilized control sample demonstrated notably higher bitterness and pungency scores compared to fertilized samples, consistent with its higher phenolic content. Importantly, all evaluated oil samples satisfied all requirements for extra virgin olive oil classification according to IOC58 standards. The classification was validated based on two main criteria. First, the median intensity of positive sensory attributes remained above zero in all samples. Second, no sensory defects were observed. These findings indicate that NPKNF fertilization may influence specific sensory attributes without compromising the oil’s classification as extra virgin olive oil.
Regarding the measurements taken during the “off-year” season, data from Table 8 reveal that foliar application of NPKNF significantly increased the number of inflorescences per shoot. The most pronounced effect was observed at a concentration of 4000 mg L−1, which yielded the highest recorded value of 8.76. In contrast, the control treatment exhibited the lowest number of inflorescences per shoot (3.53). Additionally, Table 8 demonstrates that all tested NPKNF treatments significantly enhanced flowering density compared to the control. The highest flowering density (31.73) was observed in trees treated with NPKNF at 4000 mg L−1, whereas the control recorded the lowest value (16.86). Furthermore, the data indicates that NPKNF application had a significant positive effect on the number of flowers per inflorescence. The highest number of flowers per inflorescence (13.34) was observed in response to NPKNF application at 4000 mg L−1, while the control treatment recorded the lowest value (7.72). However, with respect to the percentage of perfect flowers, no significant differences were detected among the treatments, as shown in Table 8.
Fig. 3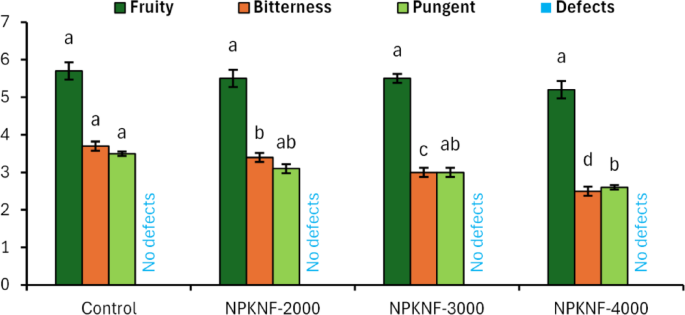
Effect of spraying NPK nano-fertilizers on sensory evaluation of ‘Picual’ olive trees during the “on-year” season. Identical letters within the same trait indicate the absence of statistically significant differences between treatments at the 5% significance level. The error bar represents the standard error (± SE).
The observed increase in flowering intensity aligns with the findings of Erel et al.16who reported that the availability of nitrogen, phosphorus, and potassium influences the number of inflorescences per shoot in olive trees. This improvement in flowering characteristics may be attributed to the role of NPK nano-fertilizers in enhancing chlorophyll content and photosynthetic activity, thereby promoting flower development75.
Table 8 Effect of spraying NPK nano-fertilizers on flowering parameters of ‘Picual’ olive trees during the “off-year” season.
The data presented in Table 9 indicate that the applied treatments did not result in statistically significant differences in the initial fruit set percentage. However, a significant and progressive increase in the final fruit set percentage was observed, with the highest recorded value of 18.6% at the highest NPKNF concentration. Furthermore, the results demonstrate significant differences among treatments in terms of fruit drop percentage. The control exhibited the highest fruit drop rate (53.31%), whereas the lowest percentage (37.01%) was recorded in trees treated with NPKNF at 4000 mg L−1. Regarding yield, Table 9 also shows a significant increase in yield (kg per tree) in response to NPKNF treatments. The highest yield (7.5 kg per tree) was obtained with the application of NPKNF at 4000 mg L−1, while the control treatment resulted in the lowest yield (3.5 kg per tree). Additionally, the data indicates that alternate bearing severity decreased significantly with increasing NPKNF concentrations. The lowest alternate bearing severity (85.4) was observed in trees treated with NPKNF at 4000 mg L−1. The results clearly demonstrate that NPKNF application enhanced the final fruit set and yield of ‘Picual’ olive trees while reducing fruit drop and improving fruit retention. These findings are consistent with those of Gad et al.75, who reported that foliar application of nano-potassium silicate significantly improved the final fruit set, yield, and fruit retention in the ‘Ewais’ mango cultivar. Similarly, Saied37 observed an increase in yield and fruit retention in mango following the application of NPKNF. Davarpanah et al.40 also reported that nano-nitrogen application in pomegranate led to increased fruit yield. The observed improvements in fruit yield and retention may be attributed to enhanced nutrient concentrations in the leaves, which have a substantial influence on fruit growth and overall productivity75,84.
Table 9 Effect of spraying NPK nano-fertilizers on fruiting parameters, yield (kg per tree) and alternate bearing severity of ‘Picual’ olive trees during the “off-year” season.
Generally, the targeted application of nano-fertilizers demonstrates considerable promise in enhancing crop performance through improved nutrient delivery and uptake. Across various crops, including wheat98,99, maize99, tomato100, and soybean101, nano-fertilizers have been associated with increased enzyme activity, enhanced photosynthetic efficiency, and more effective nutrient transport. These physiological and metabolic improvements collectively support healthier plant development and higher productivity. The observed benefits highlight the potential of nano-fertilizers as a viable strategy for advancing sustainable agricultural practices.
Furthermore, the findings indicate that NPKNF application effectively reduced alternate bearing severity. This result aligns with the studies of Kotsias et al. and Haberman et al.19,20, which reported that nitrogen fertilization mitigates the phenomenon of alternate bearing in olive trees. The observed increase in olive yield during the “off-year” and the reduction in alternate bearing severity may be due to the role of NPKNF in promoting the development of new shoots in the preceding “on-year”. Since olive trees bear fruit on one-year-old shoots, an increased number of new shoots in the “on-year” positively impacts flowering and fruiting in the subsequent “off-year”.
Dining and Cooking